[this document
is in process of assembly - l.e. 07.06.09]
Symbiosis as a term is used to refer to the character
of interactions amongst dissimilar living organisms in intimate
relation of some sort or other, specifically those between a symbiont
(guest/invader) and a host (provider/prey). As a topic people could
speak cogently about, the idea probably rose to common awareness
in the 1970’s and 80’s with the work of academic pioneers
such as Lynn Margulis and the team of Varela and Maturana. Their
research revealed startling and unexpected depths to relations that
were previously considered in a primarily mechanical light.
Though James Lovelock may have coined the term or
brought it to common awareness with his book Gaia — it
was the work of Margulis, the wife of the late Carl Sagan and mother
of Dorion Sagan — that gave proof to a startling set of postulates.
The strong implication of her research was that the concept
of competition as the basis of the evolution of species is at best
incomplete, and at worst devastatingly misfounded.
Her work and theories met with many years of ridicule,
primarily because their implications could change evolutionary theory’s
primary postulate, and shatter the existing towers of dogma
in many simultaneous domains.
In time, her research generated models demonstrating
that the evolution of the animal-cell (eukaryotes) was most probably
the result of symbiotic mergers — which were the source of
the complex cellular ecosystems that lead to animals and humans.
You could say that the ‘entity’ writing
this text, and the one reading it are more ‘cellular hyperstructures’
than they are ‘human’ or ‘animal’ —
and you’d be correct.
~ : O : ~
The ongoing reflective complexification of the available and embodied
relationships in the primordial Earth eventually led to a sort of
fusion-event — an ever-more explosive momentum was set up
in a general domain of relation that could as easily be called biocognitive
as physical. The opportunities thus potentiated led first to co-operation,
and then ever-more complex mergers.
In the modern moment, we give value-precedence to
the localized mergers, which we call animals — pretending
that the distributed mergers (ecosystems) are not in fact the same
kind of game. The truth of these matters is more startling than
either of these positions however, and requires direct experiential
contact and exploration to adequately understand.
Eventually, sub-organismal mergers (guest-in-host)
became a powerful evolutionary lens, and this generated enough opportunity
and momentum to cover the planet with millions of species of plants
and animals — countless sizes, assemblies and scales of organismal
co-evolution.
These mergers resulted in previously inconceivable spectra of cooperative
sentience and opportunity lensing. In organismal unity, previous
competitors or consortia were empowered to co-create and sustain
something like a biorelational fusion explosion: they set
up an exponentially expanding ‘standing wave’ of diversity
and complexity magnification, the result of which was new ways
of doing this more adeptly. The entire game was one of cultivating
ever-more diverse and complex connectivity, through the vehicle
of organismal relation and assembly.
These initial sparks ignited wave after wave of biological
treasure: the opportunity to overcome impossible obstacles, while
assembling ever-more relationally cogent complexity. Whether localized
in a single organism, or distributed over multiple terrains and
domains — the planet herself has been a sentient solar nursery
for This ‘by-product’ of diversity grew ever-more adaptive
with every sustainable symmetry of relation that was added in process.
Within these ‘new ways’ there were as
yet unembodied branches that would lead out of simple organism entirely
— to the locally-embodied hypersentience that was the precursor
to and is still the living source of our human consciousness: animals.
~:O:~
The meaning of this is clear: the power of competition
in nature is not as a basis or inspiration for evolution —
it is instead merely one of myriads of elements that together allow
for the diversification and prosperity of an entire organism.
Whether we look at this organism in terms of an individual, a collective,
an ecosystem or a planet — the only place competition
acts omnicidally is in human ideologies.
The source of this is not our nature, or some demon
pressing us on with whispered obligations — it is our ideas,
their shape and what they arise in real service to — which
is too often their own preservation.
The vast detriment visited upon of billions of populations
of anciently conserved and preciously elaborated sentiences by our
activity is a gesture fundamentally directed at ourselves in
every instance.
Every malformed attack on the terrestrial environment
and modern survivors vastly weakens and harms our own organism,
our human and physical relations, and the living experience of our
sentience in ways our science is not yet prepared to even glance
in the general direction of.
Consider what would happen in your own body if the
entirety of the most complex cells suddenly ‘got some new
ideas’ and began erasing every other type and instance of
organism or organismal terrain at their whim, replacing it with
what ‘they thought should be there’ — such as
machines and transports of mechanical power. Were something like
this to happen to an animal it would be dead before having had any
hope of understanding what was going on.
Luckily, Earth is slightly more resilient than this.
For the past 1000 years, at least — our planetAnimal has been
in precisely this position. I believe that the entirety
of the conserved organismal sentience of Earth stands at this moment
in the direct path of a vast variety of threats we as a species
are either inventing and nurturing, or ignoring.
In order to understand why, we will benefit from deeper
explorations into organismal purpose and relation. The meanings
we will arrive at are far more poetic than scientific: they are
living languages of meaning that value hearts before minds,
and content before structure.
~ : O : ~
guest
= self
An implication of Margulis’ work in the evolution
and meaning of endosymbionts (inward guests) is that animal cells
(eukaryotes) evolved from bacteria-like (prokaryotic) precursors
who formed intrinsic and consortial alliances. These new forms of
relation afforded participants the power to assemble synergistic
lenses capable of translinear magnifications of organismal effort,
opportunity and general prosperity.
The startling underneath of this is easily overlooked:
the historical and evolutionary path of complex animals is symbiogenic;
that it to say — elementally co-operative to the
point of assembling animals out of dissimilar animals for the sake
of mutual preservation, prosperity, and reproductive success.
If this process is what is assembling the ‘competitors’
we see through the pseudo-Darwinian lenses of our current understandings
it means that what we formally believed and acted upon as being
competition was never anything even related to what we modeled it
as.
~ : O : ~
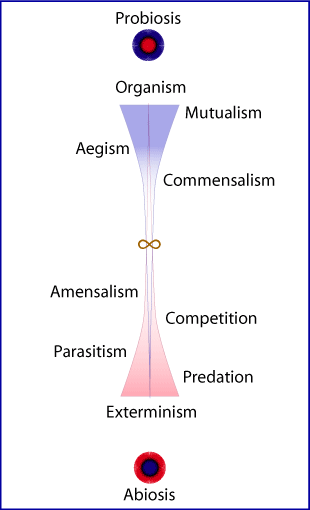
treeTree
An idealized model of the polarities of organismal relation. In
this model each primary element is comprised of sub-assemblies from
other elements, as well as circumstance (environment).
From within the perspective of any element, the map and significance
of the elements changes dramatically. It would be difficult indeed
to locate a better source or schema for the communications games
animals and humans are constantly elaborating.
~ : O : ~
Organism:
Symbiont and host function as a biocognitive and biorelational
unity. They are no longer formally discrete.
This is the physical peak of probiotic endeavor. What
was at first unified (in the beginnings of life on Earth), and then
divided (and complexified), is now re-unified to grand effect.
Symbiogenesis is the emergence of a new organism as
the result of a merger between formally dissimilar organisms.
Mutualism:
Participants enjoy mutual benefits from association
with minimal or nonexistent detriment to either.
Trophobiosis is a form of mutualism where a symbiont-provider
receives food-care from a protector-host organism.
Aegism:
The symbiont is afforded shelter, benefit or protection
by the host with no effect upon the host.
Epizoism/Ephytism:
Symbiont dwells on a surface of a host.
Endoecism:
Symbiont dwells in the host’s shelter.
Phoresy:
Symbiont utilizes the host as a vehicle of transport.
Inquilinism:
Symbiont dwells within the host, treating it as shelter. The larger
organism is generally presumed to be the host.
Metabiosis:
An indirect relation. The symbiont utilizes a product of dead hosts,
without causing harm or death to those hosts.
Commensalism:
The term comes from ancestors meaning ‘sharing
the table’ and refers to the sharing of resources (primarily
food) amongst organisms without detriment to participants due to
sharing.
Neutrality:
There is judged to be no appreciable effect because
the organisms are not in contact, or at least not in any way that
would meet our standards of qualification.
Amensalism:
The symbiont negatively impacts another neutral
organism, with no resultant gain for either.
Competition:
A relation in which the mutual presence of organismal entities is
detrimental to each other in terms of terrain needs, resource acquisition,
reproductive process, and transports related to these elements.
Parasitism:
A relationship where a presumably undesired symbiont gains benefit
from close interaction with a host who is primarily harmed in the
process.
Parasitoids: Parasites which commonly result in host
death.
Hyperparasites: Parasites which parasitize themselves,
such as a female organism who is a hunting parasite, yet is herself
parasitized by an obligate male of her own species.
Predation:
Primarily phagic (eating-motivated) relations between
predatory organisms and various or singular species of prey. The
relationship is lethal to the prey and nourishing to the predator.
The predator is commonly larger than the prey. In some forms co-operative
hunting results in wider access to prey.
Exterminism:
Functionally suicidal relations with other organisms
organized toward omnicide. The result is autophagic since any
participant is in effect a participant in a single animal.
As xenophobic modes of relation proceed into manifest activity at the
species-scale, the result is an exponentially receding access to
previously available resources, transports and relations —
particularly in terms of reproduction and cognitive opportunity.
In effect, every act of biocide is occurring in the
estranged organism’s own body, relations, lineage, children,
‘mind’, and future. Not in a ‘cause - effect’
spiral, but instantaneously.
~ : O : ~
Here are a few other terms related to our explorations
of symbiosis which it will prove helpful to be generally familiar
with:
host / guest
host / symbiont:
Usually the larger organism is considered the host,
and the smaller in the relation the guest, or symbiont. Thus an
animal cell might be modeled as the host of the organelles it exists
in obligate endosymbiosis with. From the position of the organelle
— the host has become a living universe — a sort of
penultimate ectosymbiont. We might speculatively dub this a macrosymbiont.
ecto
/ endo symbiosis:
An endosymbiont relates from within an organism. An ectosymbiont
relates with the surface(s) of a host. (on or within the surface).
Environmental symbionts such as a cat and its prey-species appear
to be outside
of these classifications, although all terrestrial organisms could
be said to be endosymbionts of Earth.
Obligate:
The relationship is required for the survival of the participant
organism(s).
Conditional:
The relationship is based on conditional factors subject to cyclic
or circumstantial change.
Permanent/Temporary:
Some forms of symbiotic relation are permanent, some
temporary. Not all the permanent forms are obligate.
Arbitrary/Circumstantial:
Most formal discussions of symbiosis would not account
these modes of relation as being of equal significance to more obviously
repetitive relations. Nonetheless, this domain forms a vast garden
of potential and momentum from which the formal relations may themselves
take their inspiration and shape.
~ : O : ~
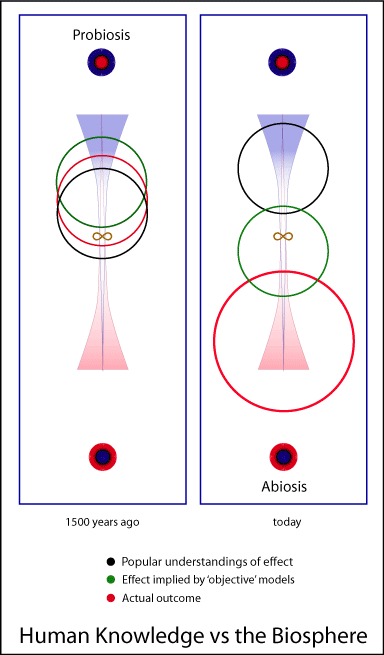
If we begin with a perspective which asserts
or implies that all individuals are discrete, we can be easily
encouraged to join the active and theoretical proponents of the ‘evolutionary
fact’ of competition as the formative basis of organismal
relation and activity.
Yet such a choice would be ill-advised for a number
of obvious reasons, perhaps foremost amongst which is that our initial
perspectives will tend to enforce themselves as fact in
our experience. This is due to the often maddening truth that the
way we perceive the terrain will tend to define the character and
adeptness of our own relations with it — thus shaping
it toward the expectations implicit in our perspective. So
too will it color our understandings and access to the real transports
and opportunities of relation, however they may be actually arrayed.
The living fact that assembles organismal sentience
at the animalian, ecological and planetary scales is a simple thing
to state: explosive magnification of relational potentials in
unity. This is the source of sentient complexity on Earth and
in our species. While the outcomes of this game may be skewed into
an interpretation that implies competition is the motivator, there
are invisible forms and modes of competition we must examine clearly
before we decide what our observations about competition mean.
The symbionts of Earth are no more in competition than the cellular
constituents of the biospheres we call our bodies — and that
competition is for the opportunity to ever-more adeptly insure survival
of the whole assembly — not for the position at the
top of some imaginary pyramid. What’s more, ‘pure competitors’
are in action and fact the enemies of the environment they arise
with(in).
~ : O : ~
The range of symbiotic behavior amongst terrestrial
organisms requires that we simplify to a degree, and also that we
pick out specific form-aspects of relation between organisms. Biology
encourages us to select the physical aspect, and thus the relations
explored involve food, shelter, transport and terrain control. When
classing a given relationship between organisms, the most generally
accurate character of the relation is what is primarily taken into
consideration. Thus a given organism may be generally predatory,
yet may also maintain a variety of non-predatory relations with
other hosts and symbionts. Of course, any complex animal is already
host and symbiont...
At the upper polarity of our tree-diagram is a place we might liken
to marriage — organism. Endosymbionts become internal elements
in the host they were once discrete from. Obligate endosymbionts
are a functionally unified organism, because neither host nor guest
can survive without each other. All animals (and many other forms
of life) are really just complexly evolved ecological vehicles existing
as the result of precisely these sorts of mergers.
The lower polarity of our model is omnicide. At this position a
given organism takes an actively abiotic stance toward lifeForms
and assemblies in general, including their own kind. Homo sapiens
is the only openly omnicidal participant in the biosphere that we
are aware of. Viruses and cancers are parasitic — but they
are a vast distance up the ladder from omnicide, which serves no
purpose but the arbitrary and hateful extermination of organismal
sentience, relation and opportunity.
Any terrestrial lifeForm represents a unique and irreplaceably
invaluable living vessel, containing and elaborating a record of
infinite conservations, mergers, histories and stories of relation.
Delicate and priceless beyond all possible exaggeration, each organism
is a living book, older and more complex than the entirety of human
technology and knowledge. This we have long known and discussed,
even if the degree to which it is true is commonly misapprehended.
The more accurate model is a seemingly incomprehensible extension
of this however: The entirety of the local organismal biosphere
is embodied in each participant completely and uniquely. Change
the container ever-so-slightly — and all participants suffer
or prosper in ongoing exponential co-reflection of the change. Remove
a single complex animal from the biosphere, and you change every
complex animal in that biosphere accordingly. The seed of this change
grows so explosively that models of it would seem absurd.
~ :O: ~
Any individual organism represents the complete and
unique organismal moment of a thread that leads all the way
back to the genesis of Life on Earth. A species
is an explosively growing library of such entities.
To understand more deeply the sources of the problems
with our species and our environment, we must reveal the understandings
that lead or encourage us to misvalue our own organismal natures,
sentience, lineages and progenitors. In doing so we will inevitably
discover that a loss in the complexity-diversity of Earth translates
instantly to a scalar magnification of that loss in human potential,
experience, consciousness and opportunity.
The reason is simple: the ‘planet’ we
call ‘Earth’ is not a planet — it’s a transentient
organismal hyperstructure. It’s thousands of billions of years
old — in every participant at every scale... not merely ‘as
one thing’. Earth is ‘more animal’ and more cognitively
miraculous than any of her constituents could possibly make accurate
models of — but we aren’t supposed to be imagining
it. We are supposed to have direct experiential access to this.
Not as metaphysics or religion: as the living fact of our animalian
birthrights, purpose, and experience. We were made for direct participation.
~ : O : ~
For thousands of billions of years, a planet-animal
of a complexity that would render all of our fictions into absurd
understatement has been assembling itself right here, with(in) its
children. The animal we call Earth is a living hypersymphony of
anciently conserved co-elaborative organismal relation. Her children
and lineages comprise a library of distributed sentience with powers
and potentials beyond the all sums of the wildest fantasies of human
technology, speculation, or knowledge.
Yet for some reason we must rapidly discover, her
first cognitively representational children cannot yet allow themselves
to admit that we ourselves are organelles in an alien sentience
that renders the sum of human science-fiction ’aliens’
entirely unimaginative in comparison.
The toyLike models and metaphors we’ve been
delivered into conceptual servitude to have so little to do with
what and where we are — as well as why. Somehow, the ways
we come to know and credential what we know are effecting a subterfuge:
they are binding us into becoming something we cannot become. In
process, they are laying waste to our human liberty, understanding,
knowledge and environment.
How do ways of knowing come to rule the entirety of
an animalian biosphere with an often omnicidal fist? Part of the
answer lies in the roots of organismal relation — and our
cursory understandings of symbiosis can be fashioned into a unique
vehicle that will allow us to more adeptly expose these questions,
their roots — and the unimagined opportunities that lie in
reframing what we believe we understand about Life, our Universe,
and ourselves.
Human
Knowledge is a ‘species’ of organismal participant in
the symbiotic basis of Earth:
Since ways of knowing result in active modes and commonly
shared or replicated lenses of relation in our species, these ‘ways
of knowing’ comprise fully functional organisms, even though
they are non-physical, and could be proven in nearly every instance
to be entirely imaginary.
Our species is unique in that ideas largely shape
the character and outcome of our organismal and conscious activity.
If the ideas are generally accurate